Quanto segue non è un calcolo di probabilità ma una serie di conti e di considerazioni volti a stabilire, tenendo conto di varie tipologie di dati, le condizioni necessarie affinché vi possa essere una probabilità non nulla di ottenere, tramite meccanismi neodarwiniani, una nuova proteina a partire da una di riferimento.
Consideriamo una proteina la cui struttura primaria sia composta da N aminoacidi. Il numero totale di proteine composte da N aminoacidi è 20N (tutte le combinazioni di parole di N lettere con un alfabeto di 20 simboli).
Se consideriamo per esempio una proteina semplicissima, composta da “solo” 200 aminoacidi, allora le proteine virtualmente possibili di pari lunghezza in tutto saranno 20200 ≈ 10260.
Assumiamo che a causa di vincoli di varia natura (fisici, chimici, biologici e ogni loro combinazione) non tutti gli N aminoacidi di una proteina siano effettivamente suscettibili di essere sostituiti tramite una mutazione, per cui indicheremo con M gli aminoacidi bloccati da tali vincoli (per brevità in seguito diremo che abbiamo M vincoli), mentre definiamo G = N – M il vero numero di aminoacidi modificabili. Abbiamo scelto la lettera G per richiamare il concetto di “grado di libertà”, per cui G sono i gradi di libertà del nostro sistema (la proteina).
Prima condizione (G massimo)
Siccome il caso in cui M = 0 e G = N implicherebbe un numero abnorme di combinazioni (come appena detto nel caso di N = 200), l’insieme di possibilità che possono essere esplorate dalle mutazioni casuali non potrebbero mai essere 20N perché ciò richiederebbe tempi proibitivi per poter avere qualche possibilità di ottenere una nuova proteina. Chiamiamo l’insieme di tutte queste possibilità “spazio delle fasi totale”, e indichiamolo con STOT = 20N.
Grazie ai vincoli, lo spazio delle fasi effettivo, che chiameremo “spazio delle fasi accessibile”, indicandolo con SA, è in genere molto più piccolo: poniamo dunque per definizione SA = 20G.
Nel caso dell’evoluzione sulla Terra, un pianeta che esiste da un tempo dell’ordine di 1018 secondi e che è composto da 1050 atomi, per stabilire la fattibilità di tale meccanismo occorre contare tutte le possibili reazioni chimiche che si sono succedute. Nell’ipotesi fortissima che si consideri come durata minima per una reazione (al di sotto della quale non si può scendere in base alla vigenti teorie fisiche) il tempuscolo di Planck pari a 10-43 secondi, allora il numero totale di reazioni avvenute nel nostro pianeta è dell’ordine di 10111. Se si facessero altre ipotesi su tale numero (in particolare più realistiche), col risultato che in generale esso sia dell’ordine di 10P, allora chiamiamo E il corrispondente esponente da dare al numero 20 per ottenere 10P (cioè 10P = 20E). Esplicitamente, E = P/Log20 (dove Log20 è il logaritmo in base 10 di 20).
In sintesi, affinché sia fisicamente realizzabile un’evoluzione neodarwiniana di una proteina deve essere soddisfatta la seguente condizione:
G < E
In altre parole, i vincoli devono essere opportunamente numerosi, cioè gli aminoacidi modificabili opportunamente pochi, per non rendere di fatto impossibile tal tipo di evoluzione. Nel caso di P = 111, E = 85.
Seconda condizione (G massimo):
Una volta stabilito che il vero spazio da considerare sia SA e non STOT, anche se SA fosse opportunamente piccolo, affinché avvenga l’evoluzione in tale spazio ridotto deve poter esistere una combinazione di aminoacidi che sia in grado di essere utilizzata dall’organismo in esame, in particolare deve dare un contributo positivo nel contesto della selezione naturale. In SA, quindi, deve esserci una nuova proteina che almeno in potenza, oltre che soddisfare gli M vincoli, sia effettivamente funzionante.
A priori, non sappiamo in tutto lo spazio STOT quante proteine di questo tipo esistano; indichiamo quindi con g il loro numero e facciamo un’ipotesi di lavoro: detta d la densità di proteine potenzialmente funzionanti in tutto STOT, assumiamo che tale densità sia uniforme in STOT. Tale assunzione corrisponde alla seguente formula: d = g/STOT
Stiamo quindi assumendo che le proteine capaci realmente di funzionare abbiano la stessa concentrazione andando ad esplorare zone diverse dello spazio STOT. La grandezza g è stata chiamata così perché, nell’ipotesi di densità uniforme, sta a quantificare il “gradualismo”: quanto più le proteine funzionanti sono numerose, cioè quanto maggiore sono d e g, tanto più, data una proteina funzionante, ne si può trovare una “vicina” (ottenuta cioè cambiando pochi aminoacidi) che sia a sua volta funzionante.
Prima osservazione: se anche all’interno di SA le varie combinazioni avessero probabilità diversa, non cambierebbe il nostro discorso, perché sarebbe come lavorare con un G “un po’ più piccolo” del caso in cui tutte le combinazioni di SA siano equiprobabili (a rigore G, come anche N e M, è un numero intero positivo, ma mi si perdoni la licenza poetica).
Seconda osservazione: se si volesse affrontare il caso di una proteina che cambia perché passa dall’avere N aminoacidi a (N+1) aminoacidi, vale ancora lo stesso discorso come se fossimo partiti da una proteina con (N+1) aminoacidi e G almeno pari ad 1.
Riprendendo il nostro ragionamento, per rendere fattibile un evoluzione neodarwiniana completa di vincoli dentro lo spazio delle fasi ridotto SA deve esserci almeno una proteina funzionante tra tutte le g proteine di questo tipo; nell’ipotesi di densità uniforme, tale condizione si esprime dicendo che SA x d ≥ 1. Di conseguenza, in base alle definizioni di SA e di g, la disuguaglianza corrisponde alla seguente formula: 20G x g/20N ≥ 1 cioè
G ≥ N – log20g
(in questo caso compare al secondo membro il logaritmo in base 20 di g).
In altre parole, quanto più G è grande, tanto più abbiamo bisogno di un gradualismo molto spinto (g grande), perché per definizione N > G.
Condizione generale e discussione
Riassumendo quanto detto nei due paragrafi precedenti, la condizione che deve soddisfare la grandezza G, cioè il numero realmente modificabile di aminoacidi in seguito all’applicazione dei vincoli è la seguente:
N – log20g ≤ G < E
G non può essere troppo grande, perché diventerebbe impraticabile per ragioni di tempo e di materia la ricerca per mutazione e selezione di una forma evoluta della proteina di partenza; ma G non può nemmeno essere troppo piccolo, perché i vincoli rischierebbero di escludere dallo spazio delle fasi ridotto un tipo di proteina funzionante (in potenza).
Purtroppo, l’esponente E nella realtà è sicuramente ben più piccolo di 85, perché possiamo quasi certamente considerare esageratamente breve l’intervallo di tempo del tempuscolo di Planck, così come non ha senso prendere in considerazione tutti gli atomi della Terra dal momento che la vita è possibile per definizione solo nella biosfera (e non nel mantello e nel nucleo). Infine, spesso solo in tempi molto brevi rispetto all’età del nostro pianeta è stato possibile il realizzarsi di un certo caso di evoluzione.
D’altro canto, si potrebbe risolvere tale limite teorico assumendo che in realtà basterebbe partire da un G piccolo, per poi immaginare che la nuova proteina, diversa per struttura primaria solo per pochi aminoacidi, sia soggetta però ad un nuovo set di vincoli tali che il “nuovo G” sarà dello stesso valore ma riferito nello specifico ad altri G aminoacidi (diversi dai precedenti).
Così facendo, però, dovremmo assumere che sia quindi realizzabile un’evoluzione tramite successive iterazioni di cicli formati da piccole variazioni sostenute dai vincoli e poi dalla selezione naturale, cioè da un forte gradualismo (g grande). Tale assunzione sarebbe tanto più necessaria quanto più la proteina di partenza sia grande (N grande), dal momento che, a causa della condizione di massimo, G non può essere troppo grande (cioè G deve essere molto minore di N).
Sappiamo che la storia evolutiva in realtà ha avuto varie fasi di “salti evolutivi”, di divergenze o singolarità, per cui in base ai dati attuali, probabilmente g non lo si può assumere grande.
Le principali soluzioni che riteniamo di presentare per tale paradosso sono due:
1) esistono e sono esistite molte più proteine (e quindi tessuti, organi e specie viventi) di quelle attualmente note e paleontologicamente registrate. Se così fosse, allora il parametro g sarebbe solo apparentemente piccolo (il gradualismo sarebbe quindi solo nascosto da lacune nei dati). Tale spiegazione sarebbe però epistemologicamente ambigua, perché si farebbe ricorso ad una spiegazione di assenza di dati invece che ad un cambiamento di ipotesi, con fortissima violazione del principio di Occam. Inoltre, dovrebbe essere corroborata con un numero massiccio di nuove proteine sconosciute, a causa dell’andamento logaritmico della condizione di minimo.
2) è falsa l’ipotesi di densità uniforme in STOT delle g proteine. In tal caso, il rispetto delle due condizioni sarebbe soddisfatto postulando una densità non più uniforme ma diradata in certe zone di STOT e altamente concentrata in altre. Benché tale ipotesi sia molto più plausibile della prima, avrebbe un’implicazione importante: se la densità d delle proteine potenzialmente funzionanti si concentra proprio nelle aree SA, vuol dire che i vincoli che sono stati artefici della limitazione di STOT sono a priori a favore della fitness, avremmo quindi a che fare con vincoli dipendenti dalla fitness e in modo positivo. Se tali vincoli sono quindi ciò che fa collassare l’immenso STOT in SA (basti pensare che l’applicazione di anche un solo vincolo fa in modo che SA sia solo il 5% di STOT, essendo 20N-1/20N = 20-1 = 1/20 = 0,05), allora il ruolo della selezione naturale sarebbe infinitesimo, così come quello del caso (inteso come meccanismo indipendente dalla fitness). In altre parole, staremmo trattando una spiegazione non-darwiniana dell’evoluzione mutuamente esclusiva a quella neodarwiniana.
Conclusioni
Per testare tale modello alla base dei criteri di realizzabilità, occorrerebbe un software che abbia in memoria e analizzi i dati delle proteine note. In particolare si dovrebbe partire dal valore di N più basso, contare tutte le proteine aventi la stessa lunghezza (stesso N) ottenendo quindi un valore sperimentale di g. Per ogni N, si calcolerebbe poi N – log20g e lo si confronterebbe con G: se G risulta maggiore, occorre poi confrontare G con una stima meno approssimativa di E. Nel caso non venisse soddisfatto uno dei due criteri, occorrerebbe affrontare le questioni sollevate alla fine del paragrafo precedente.
.
.
.
55 commenti
La spiegazione di come potrebbe essere possibile la formazione di una goccia nell’oceano senza preoccuparsi dell’oceano…
Direi proprio il contrario… visto che i “vincoli” che favoriscono la fitness rappresentano non solo l’oceano, ma anche il cielo e la terra
Intendevo semplicemente dire che anche qualora si dimostrasse la probabilità non nulla e matematicamente significativa di sintesi “casuale” da una proteina A ad una proteina B non si sarebbe spiegata di certo la complessità a tutti i livelli di osservazione (ambientale, ecologico etc.) di tutto ciò che possiamo osservare nel mondo organico che necessiterebbe ancora di una spiegazione e di um modello esaustivo…
Non volevo dire che lo studio di Htagliato nella finalità di raggiungere il suo obbiettivo non ne abbia tenuto conto, ma solo che quest’esercizio a mio parere non è di grande utilità.
Insomma, a mio parere HTagliato ha speso il suo tempo per contrastare un “problema” che non ha implicazioni così enormi ed importanti paragonato al mondo organico nella suo complessità e totalità.
Bel lavoro, HT.
Io vedo come unica, scientificamente accettabile, soluzione del paradosso, la seconda opzione, quella in cui la selezione naturale ha peso “infinitesimo” (più esattamente nullo, come vale per ogni tautologia) e in cui d si concentra in SA. Non sarebbe la prima evidenza in scienza naturale di fine tuning antropico…
Chiamare “ambigua” dal punto di vista epistemologico la prima soluzione al paradosso mi pare un eufemismo molto politicamente corretto. Stai preparandone un articolo scientifico?! Con l’aiuto di un biologo-statistico magari, che ti possa confortare con il test del modello…
Piacere rivedere la firma di HT su queste pagine. Non riesco però a cogliere il senso e l’effettiva utilità di questo modello, a mio avviso troppo teorico, in quanto non riesco a cogliere una linea di continuità concreta con il quadro storico-biologico completo. Non riesco a capire se si voglia in qualche modo coinvolgere anche il problema della proteinosintesi primordiale, o semplicemente analizzare soltanto la possibilità di una trasformazione di una proteina già esistente in un’altra. Perché se parliamo della prima opzione allora ci sarebbe secondo me da coinvolgere quantomeno anche il contesto ambientale e organico che si presume possa essere stato espresso nella Terra di qualche miliardo di anni fa, e quindi sostanzialmente affrontare il problema complesso dell’abiogenesi (fenomeno scientifico comunque avvenuto), se invece parliamo della seconda, come sembrerebbe invece suggerire il sottotitolo, allora non vedo dove sta il problema. Il processo del cambiamento di una proteina ad un’altra è un fenomeno comunissimo, che si verifica frequentemente e che, in generale, coinvolge tutto l’apparato sintetico a partire dalla mutazione genetica e passando per m-rna, ribosoma, codone. Come per esempio accade per l’anemia falciforme, in cui una sola sostituzione basica da adenina a timina (per mutazione puntiforme) determina la sostituzione dell’aminoacido acido glutammico con valina. E tutti conosciamo i motivi per cui l’allele è poi conservato grazie al vantaggio neodarwiniano dell’eterozigosi in termini di resistenza alla malaria. Oppure i casi dei geni regolatori, es. FOXP2 e la sua implicazione sul linguaggio umano o HERC2 per gli occhi azzurri. Nel caso di altri tipi di mutazione o per esempio di frame-shift le conseguenze in termini di sintesi proteica ed espressione fenotipica, possono anche essere di maggior rilievo. Quindi, ripeto e mi scuso per l’eventuale mia sbagliata interpretazione, qual è il problema?
Secondo me, dott. Vomiero, l’abiogenesi non è “un fenomeno scientifico comunque avvenuto”, perché non abbiamo una testimonianza storica che sia avvenuto né un modello scientifico di come possa essere avvenuto, ma è soltanto una congettura scientifica. Ci sono anche scienziati, tra cui biologi e fisici, che lo ritengono semplicemente un assioma, cioè un postulato indimostrabile.
Ha ragione prof.Masiero, il termine fenomeno è perlopiù legato a qualcosa di osservabile. Intendo, tolta la Creazione, che non è un concetto scientifico, la congettura più plausibile all’interno dell’ambito scientifico, non necessariamente sulla Terra, ma comunque da qualche parte nel nostro universo.
Anch’io la ritengo la congettura più plausibile, ed anche forse un giorno risolvibile (mentre non considero le facoltà intellettive e la libertà umane esplicabili scientificamente). Devo osservare però che anche “plausibile” non pertiene alla scienza naturale, ma al gusto personale. Ci sono scienziati, come Bohr, che assumono l’abiogenesi come postulato, quasi in una sorta di dualismo tra materia inanimata e materia animata o di vitalismo o comunque di “autonomia della biologia” (E. Mayr); altri che non distinguono tra materia inanimata e animata e quindi, in un continuum, negano la necessità stessa dell’abiogenesi; altri ancora che ricorrono alla panspermia postulando un principio vitalistico alieno; ecc., ecc.
“forse un giorno risolvibile”
Quindi anche FORSE NO.
Stavo riflettendo prof.Masiero sul fatto che, come dice lei, il concetto di plausibilità possa essere anch’esso una questione di gusti. Io credo che questa affermazione possa essere tranquillamente accettabile sul piano socio-culturale che contraddistingue ogni persona dall’altra, ognuno di noi infatti possiede un filtro cognitivo proprio che è più o meno diverso da quello di una qualsiasi altra persona. Non sono molto d’accordo invece se parliamo di scienza. Il processo scientifico infatti è, a mio avviso, per le modalità stesse con cui produce conoscenza, un “sistema cognitivo” che, più di ogni altro, ci fa avvicinare al concetto di oggettività. All’interno dei propri confini e dei propri limiti, ogni descrizione scientifica del mondo è, di fatto, in un preciso momento storico, la più plausibile, per usare una felice espressione di Calvino è la (più affidabile) “concrezione del mondo”. La questione sollevata da Voce nel deserto in merito a eventuali posizioni influenzate da condizioni di ateismo o altro, credo invece non abbia molto a che fare con la scienza, o quantomeno non è (per la scienza) di nessuna utilità pratica e funzionale.
Se gli scienziati nelle loro ricerche, dott. Vomiero, si muovessero sempre lungo le linee che la maggioranza di loro ritiene intersoggettivamente più plausibili (non oggettivamente, perché la plausibilità non è misurabile!) la scienza, o piuttosto la tecnica, procederebbe quietamente in avanti, ma senza quei grandi cambi di paradigma nelle teorie che costituiscono, secondo me, l’aspetto più affascinante ed epistemicamente più rilevante della scienza naturale.
La storia c’insegna che le grandi scoperte in scienza consistono sempre di confutazioni delle teorie correnti, considerate fino ad allora plausibili; scoperte grandi proprio per la loro discontinuità e che possono essere fatte solo da ricercatori giovani e rivoluzionari.
Perchè non tiene conto di come la posizione esistenziale di uno scienziato possa influire sull’interpretazione dei dati oggettivi e dei criteri di indagine ricerca…
Il suo/vostro concetto di “scienza” è metodologicamente ateo e per questo limitato e se Dio esistesse girereste in tondo all’infinito producendo teorie false in continuazione perdendo tempo.
Buona “scienza”…
Sì, io credo a quella favoletta. Non conosco il suo background, Voce, ma guardi che la scienza e lo scienziato sono ben formati e abilitati a gestire il principio di “non sovrapposizione dei magisteri”. Se uno fa scienza, fa scienza e basta e quando lavora, lascia inevitabilmente fuori dalla porta il suo bagaglio di eventuali “credenze” extrascientifiche. Le opzioni Creazione, Dio, ID o quant’altro, possono anche essere interessanti e discutibili sul piano personale, ma dal punto di vista della scienza non fanno fare nessun passo in avanti, è questo il grande problema. Guardi, ho un esempio che conosco bene e sono io, anche se le potrà sembrare strano. Io ho il mio credo, Voce, io prego e vado a Messa, credo in un Qualcosa, anche se non so dire esattamente in cosa, io non ho la presunzione di dire che cosa sia la Rivelazione, o la Verità, non ne ho gli strumenti e come me, chiunque altro, non ci sono esseri “superiori” terreni che abbiano la Verità in tasca, tantomeno gli scienziati. E’ per questo che guardo con sospetto ogni tipo di Fede fondamentalista e ogni tipo di mito, e apprezzo così convintamente la grande rivoluzione concettuale portata finalmente dalla scienza moderna, dopo millenni di mitologia.
Quand’ho usato il termine “Dio” ho post-posto la parentesi “concetto da chiarire“.
Non era mia intenzione entrare in questo argomento (infinito), ma le dico solo che non mi riferivo nè al concetto di “rivelazione” nè tanto meno ad un concetto personale di “veritá“.
Alludevo al fatto di contemplare l’eventualitá della singolarità sovrannaturale come causa accettabile laddove, dopo aver risalito la china delle cause naturali possibili alla ricerca dell’origine delle catena completa delle cause stesse (cioè arrivati alla causa prima) si arrivi ad punto nel quale le leggi naturali non sono in grado di consentire la prosecuzione della ricerca stessa e un sua ipotetica postulazione potrebbe spiegare in maniera esaustiva e semplice i fenomeni OSSERVABILI ED OSSERVATI (che devono quindi rimanere il PUNTO DI PARTENZA DELLA RICERCA) i quali, alternativamente, per giustificare la loro esistenza, necessiterebbero della formulazione di speculazioni ultra-complesse e “stravaganti” (per usare un eufemismo) spacciate per “teorie scientifiche” laddove in realtà non sarebbero nient’altro che speculazioni non verificabili ed assurde spacciandole pure come “ragionevoli”…
Le faccio un esempio concreto solo per rendere più chiaro e meno fumoso il concetto, iper-semplificando ed ignorando volutamente i dettagli tecnici per astrarre il tutto solo a livello concettuale: diciamo che venisse postulato un modello per l’origine del sistema solare, estensibile anche tutti gli altri sistemi astronomici osservabili nell’universo, secondo il quale tutti gli astri del sistema solare sarebbero fuori-usciti da un unico astro originale (chiamiamolo “sole originale” S) attraverso la sua messa in rotazione ad una velocitá critica (diciamo R) ben precisa e calcolabile sulla base di leggi fisiche note, verificabili e ripetibili (tralascio volutamente i dettagli), velocità di rotazione la quale, sempre in base a leggi fisiche ed esperimenti effettuati e ripetibili è in grado di produrre l’espulsione dal corpo originale di materia la quale otterrebbe così in certa massa, moto di rotazione etc. e che, in concomitanza con altre forze come quella gravitazionale, assumerebbe una certa orbita nello spazio attorno al corpo originale andando insomma a spiegare in maniera esaustiva l’origine del sistema solare (ma non solo).
R però è maggiore di r (la velocità di rotazione attuale del sole) il che implicherebbe che S ad un certo punto nel tempo avrebbe dovuto o accelerare la sua velocità di rotazione (se già in rotazione) o “mettersi in moto di rotazione accelerato” (se da immobile) e che dopo l’espulsione dell’ultimo corpo da S il sole attuale rimanente (s) avrebbe dovuto decelerare la propria velocità di rotazione raggiungendo r.
Insomma, diciamo che forza centrifuga, centripeta e rotazione sarebbero la chiave di volta per comprendere l’origine dei sistemi astronomici osservabili nell’universo (semplificando).
Lei (e chi ragiona come lei) mi farebbe notare che non esiste una legge naturale/fisica in grado di giustificare la singolarità che il S abbia potuto accelerare o raggiungere da immobile la sua velocità di rotazione fino ad R e che quindi, secondo l’attuale (e a mio parere viziatissimo) concetto di “scienza“, non sarebbe una teoria “scientifica” e di conseguenza si rifiuterebbe di tenerla anche solo in considerazione.
Infatti in un tale scenario l’unica causa dell’accelerazione ad R sarebbe che una “forza di origine sovrannaturale” abbia messo in rotazione accelerata S più volte producendo le conseguenze che possiamo osservare (i pianeti con le loro orbite, distanze, masse, rotazioni etc.).
Quale sarebbe la conseguenza del suo rifiuto?
Che ci si opporrebbe al suo insegnamento nelle scuole ACCANTO ALLE ALTRE di tipo naturalistico e che si obbligherebbe a cercare altre spiegazioni naturalistiche più esaustive di quelle attuali (non esaustive).
Se però le cose fossero andate proprio come ho postulato ipotizzato la conseguenza potenziale sarebbe di rincorrere all’infinito spiegazioni inadeguate, in realtà false, non esaustive e potenzialmente sempre più complesse rese “sostenibili” solo da esercizi matematici funambolici ed iper-complessi accessibili solo ad iper-esperti in materia (con le relative implicazioni) provocando di fatto e alla lunga uno spreco di risorse inutile e virtualmente infinito.
Accettando invece l’ipotesi della singolarità sovrannaturale, alle CONDIZIONI POSTE ALL’INIZIO del discorso, il “passo avanti” verrebbe fatto eccome perchè si otterrebbe un modello esaustivo per spiegare l’origine del sistema solare (e non solo) il che eviterebbe di rincorrere all’infinito ipotesi anti-logiche, funamboliche e false.
E non mi venga a parlare di “God of the gaps” perchè le condizioni che ho posto all’inizio lo escludono e una singolarità con un’ipotetica e fantomatica legge naturale sconosciuta, perchè una legge per definizione agisce costantemente e non per singolarità irripetibili.
Concludo inoltre dicendole che lei come “Cattolico” (non può che considerarsi tale visto che afferma di “andare a messa“) ha dei seri problemi sui quali dovrebbe riflettere seriamente oppure smettere all’istante di “andare a messa” per non offendere la fede che lei afferma di praticare e di seguire…
Riconoscere l’esistenza di una Rivelazione non è sintomo di “presunzione“, è una necessità logica che si impone nel momento stesso nel quale afferma di credere nell’esistenza di un’entità trascendente, personale e intelligente e quindi dotata della capacità di rivelarsi.
Trattasi bensì di FEDE e come Cattolico lei DEVE aderire ad una certa dottrina tramandata per successione apostolica del tizio nel quale nel afferma di credere andando a messa, tramandazione che le insegna che “Dio” (l’entità di cui sopra) si è rivelato e che tale Rivelazione è contenuta nella Scriptura e nella Traditio e che è proprio sulla base di quella Rivelazione che si deve fondare il motivo stesso per il quale lei dovrebbe “andare a messa“, e cioè la celebrazione dell’Eucarestia, cioè il “mistero” rivelatole da quella stessa Rivelazione che lei afferma di non sapere cosa sia e che sia “presuntuoso dire cosa sia“…
Davvero lo dico per lei: ci rifletta bene o smetta di andare a messa per rispetto di chi ci va con cognizione di causa e dell’entità che teoricamente verrebbe in quel luogo adorata…
Voce, la serie di causalità per accidente può essere infinita. Qundi si può concepire un universo che non abbia inizio né fine, insegna San Tommaso.
Nel tuo ragionamento fai ingiustificatamente un salto qualitativo tra il ragionamento scientifico che è intrinsecamente incapace di risalire alle cause sostanziali poprio per sua natura e quello filosofico che è il solo atto a considerare questa problematica nel mondo reale.
Io non ho detto che non è possibile concepire un universo che non abbia inizio ne fine (il che comunque lo sarebbe solo perchè Dio è eterno e non per altro…).
Io ho posto come condizione iniziale del mio ragionamento il caso limite nel quale una certa singolaritá oer poter essere contemplata debba per forza essere di origine extra-ordinaria /sovra-naturale perchè non spiegabile tramite le leggi naturali.
Inoltre non faccio nessun salto qualitativo: io contesto la concezione stessa che lei abbraccia di “scienza”.
“Più plausibile” per una mente atea o di un “credente” che si illude di esserlo ma che nel concreto assume comportamenti ed impostazioni atee auto-negando di fatto la sua professione di fede senza magari neppure rendersene conto…
Il problema che lei descrive è ampiamente discusso in letteratura e credo sia doveroso prima di porre domande su di un argomento informarsi scrupolosamente su quali sono le risposte che altri esperti autorevoli hanno dato sulla base delle loro nozioni.
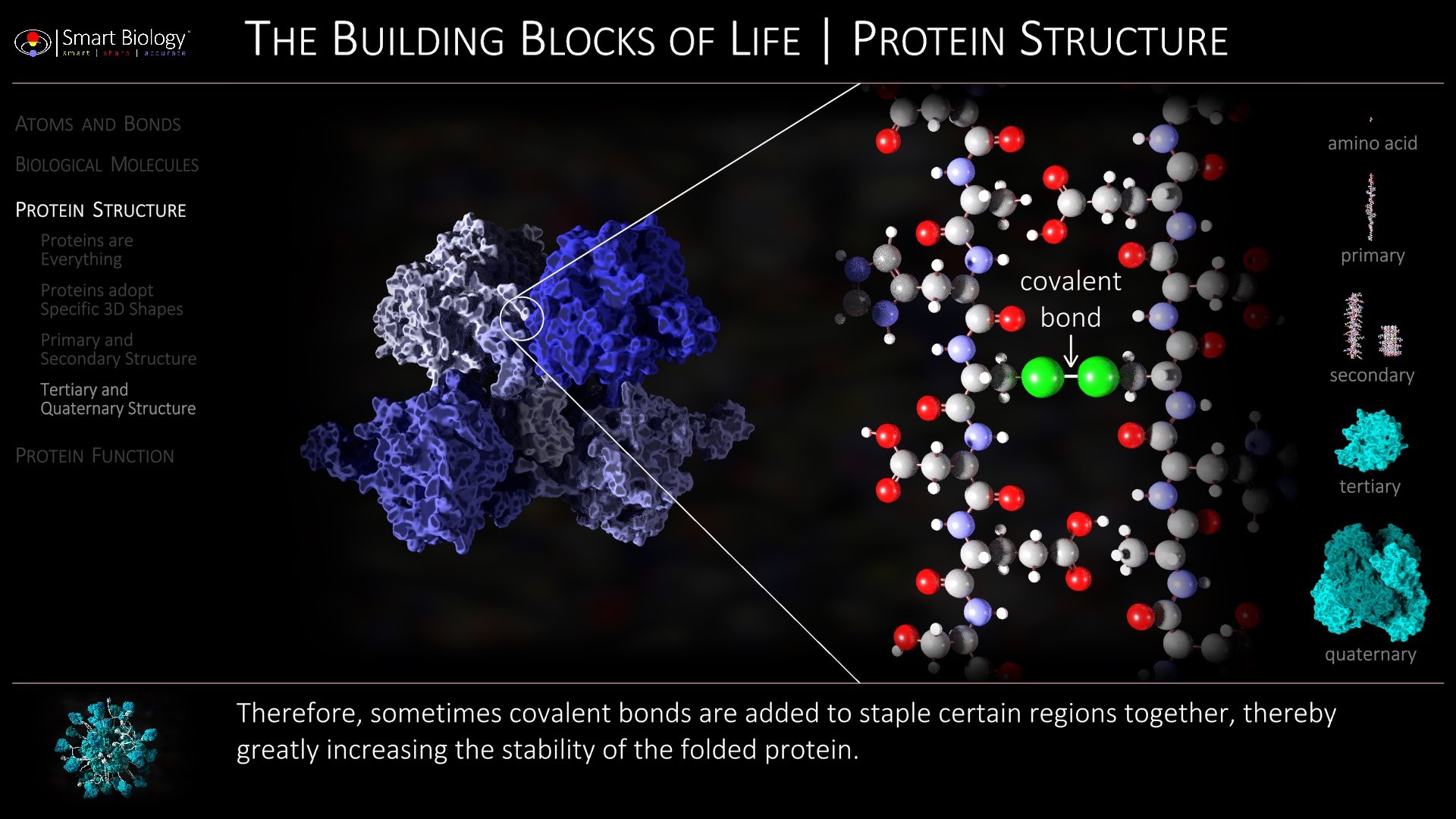
Per cominciare un appunto sul concetto che lei da di proteina “semplicissima”. La invito a consultare un lavoro sulla lunghezza media di una proteina, in particolare di quelle più evoluzionisticamente “recenti”, come ad esempio quello pubblicato su Genome Biology qualche anno fa (2010) da Capra JA et al. intitolato “Novel genes exhibit distinct patterns of function acquisition and network integration” 200AA è una lunghezza media per una proteina “recente”, con il 25esimo percentile più intorno a 100AA (proteine “corte”) e quelle “molto corte” (semplicissime come dice lei), nettamente sotto ai 100AA
Oltre alla lunghezza, al suo modello manca una sufficiente conoscenza dell’oggetto che sta modellando. Conoscendo la biochimica assumere che lo spazio delle proteine ipoteticamente funzionante sia omogeneo non è ragionevole. Le proteine infatti presentano conservazione diversa per zone diverse e maggiore è la conservazione, minore è la variabilità del sito. Quindi occorre modellare lo spazio delle possibili mutazioni in modo non lineare, cosa che riduce il numero di proteine valutabili e non credo che questo sia “un po’ più piccolo” perchè la differenza non è appunto lineare. (v. ad esempio il lavoro di Laurent Duret “Determinants of Substitution Rates in Mammalian Genes: Expression Pattern Affects Selection Intensity but Not Mutation Rate” pubblicato nel 2000 su Molecular Biology and Evolution
A tale nozione possiamo aggiungere che molte proteine non si evolvono per mera aggiunta di un aminoacido alla volta, ma per fusione di geni o pseudogeni che possono rappresentare domini (ovvero porzioni funzionali della proteina) interi, “ereditati” nella ricombinazione genica. Il modello evolutivo è quindi molto più complesso di quello che lei delinea, molto più dipendente dallo stato pregresso, che rappresenta un vincolo molto forte. Se infatti lei prende la strutture cristallografiche disponibili fino ad ora e analizza un sito di domini funzionali e famiglie proteiche come PFAM, si accorgerà che tale collezione è ben lungi dall’essere enorme e pure altamente variabile, ma presenta collezioni di domini notevolmente diverse tra loro ma internamente piuttosto omogenee. Anche questa osservazione, che segue da una opportuna conoscenza della biochimica proteica, dovrebbe condurre il ricercatore coscienzioso verso ipotesi adeguate.
Un “vecchio lavoro” in merito è del 1992: “Environment-specific amino acid substitution tables: tertiary templates and prediction of protein folds.” Su Protein Science da parte di Overington J et al. Qualcosa di più recente lo trova su Molecular Biology and Evolution del 2004 in ” Prediction of site-specific amino acid distributions and limits of divergent evolutionary changes in protein sequences.” di Markus Porto et al.
Delle ipotesi che lei fa per cercare quindi di spiegare il paradosso che ha creato la prima è poco probabile, non ci sono ragioni per pensare che “esistono e sono esistite molte più proteine (e quindi tessuti, organi e specie viventi) di quelle attualmente note e paleontologicamente registrate”. Aggiungo che l’idea di accostare paleontologia e biochimica delle proteine fa più Jurassic Park che ipotesi scientifica generale sulle proteine.
La seconda è più interessante. Se assumiamo che le proteine odierne sono “frutto” delle proteine molto semplici (ragionevolmente qualche dozzina di AA) e che da esse hanno ricevuto vincoli essenziali, quelli sono i vincoli che limitano le proteine odierne. Nessuno sa se sia ipoteticamente possibile costruire “super proteine” (per le necessità odierne) molto più efficienti di quelle attuali (e.g. con parametri enzimatici più preformanti) ed è possibile che le proteine che abbiamo oggi sia il “meglio che si può fare con i vincoli” e la selezione avviene solo su quelle, tenendo conto del fatto che sia lo spazio delle proteine, che la sequenza di ogni singola proteina, non possono essere modellati in modo omogeneo rispetto alla variabilità e al suo significato funzionale.
Forse può giovarle ragionare in termini più macroscopici. Quanti modi ci sono di organizzare le cellule di un organismo medio? Il numero è ovviamente molto più grande di quelli di cui lei tratta nel suo testo, con miliardi di cellule. Modellare la probabilità di esistenza di una specie in questo modo porterebbe a concludere che il numero di vincoli deve essere enorme, e in effetti è così. Il genoma pone vincoli ed esso stesso non è “creato” da nulla ad ogni generazione, ma varia molto poco da una versione precedente. Se oggi osserviamo certe caratteristiche (arti con 4-5 dita, colonna vertebrale, scheletro di chitina, branchie, produzione di sperma, foglie, ecc.) non è detto che siano il modo migliore in assoluto per svolgere quella funzione, ma quello residuo frutto di una serie di eventi pregressi, che hanno introdotto una miriade di vincoli che non permettono di “saltare” a un organismo a sangue caldo che fa anche fotosintesi, nonostante tali processi siano osservabili. Gli organismi infatti fanno parte di un ecosistema, si influenza a vicenda e le loro funzioni fisiologiche si influenzano similmente a vicenda, introducendo un numero di vincoli molto grande, che impedisce di osservare tutti gli organismi che ci potremmo immaginare.
Se usa questa analogia dovrebbe subito apparirle evidente che ancor prima di cercare di fare speculazioni evolutive su di un modello, dovrebbe essere sicuro di non aver perso per strada alcun elemento oggi noto utile alla modellazione, come mi pare il suo caso. Ha considerato di continuare il suo percorso di studi in biologia? Le sarebbe utile, considerata la multidisciplinarità dei suoi testi e la monodisciplinarità della sua formazione in fisica.
Gentile Nando, grazie per questa lunga ed accurata analisi, e per i tanti riferimenti agli studi già condotti su questo argomento.
Prego, si figuri. I tre riferimenti sull’argomento che ho fatto sono ben poca cosa; ciascuno di essi ha una nutrita bibliografia per le decine di riferimenti fatti.
E’ importante costruire su quanto esistente e a maggior ragione quando si dissente, citare le fonti e chiarirne i motivi, per permettere al lettore di apprezzare l’eventuale cambiamento, contestualizzandone la novità.
Sono d’accordo con lei sulla necessità di citare quanto è già stato fatto in un determinato ambito, specie quando si porta avanti una critica che vuole essere costruttiva.
Le faccio una domanda: sbaglio o nel post si parte dal presupposto che il numero massimo di reazioni possibili nella storia della Terra è calcolato considerando queste reazioni in serie, e non in parallelo come dovrebbe essere?
E’ così, ma conta poco. Anche considerando tutte le cellule mai esistite sul pianeta non ne uscirebbe un numero sensato, ed è ovvio.
Infatti è il presupposto di indipendenza degli eventi ad essere errato e quindi quel numero non ha senso nemmeno calcolarlo così.
Ogni evento biologico è presupposto per quello successivo e lo condiziona, così che non ha senso considerare tutte le possibilità in quel modo.
E’ come dire che siccome il numero di possibili posizioni delle foglie di un albero è enorme, l’albero non cresce in modo casuale ma la sua forma definitiva deve essere già nota a priori. Così di dimentica del ruolo essenziale dell’evento germinativo, di ogni singola ramificazione, e della complessità dei fattori che la determinano, vincolando la posizione di tutte quelle successive.
Con un albero possiamo ripercorrere questi eventi e nessuno si mette a dire che un albero è un miracolo. Con l’evoluzione il passato è meno intellegibile, ed è per questo che piuttosto di sforzarci di trovare ragioni per non investigarlo, dovremmo trovarne per capirlo meglio.
Grazie per la risposta.
Quindi, riassumendo, si può dire che il calcolo proposto non è corretto perché a) considera le reazioni indipendenti; b) considera una sola sequenza di reazioni in serie; c) non considera la complessità del sistema?
Più che correttezza parlerei di adeguatezza. In fondo stiamo cercando una modellazione per capire meglio. Solo un modello perfettamente predittivo sarebbe corretto e non lo sono nemmeno i modelli teorici di leggi fisiche, quindi sarebbe utopistico cercare la correttezza.
Più il modello tiene conto dei vincoli, più sarà adeguato. Il modello proposto non tiene conto di quasi nulla e ignora moltissime nozioni di biologia e biochimica di base, quindi resta molto speculativo e in ultima analisi un esercizio intellettuale superficiale, avulso dalla realtà di cui non tiene infatti conto.
Giusta la sua precisazione sulla correttezza. Vedo che condividiamo le stesse perplessità, spero che Htagliato vorrà darci alcuni chiarimenti.
Divertente il teatrino tra il professore e lo studente, ma il “presupposto d’indipendenza”, Nando, lei dove l’ha letto?
Htagliato le risponderà se crederà, io ho letto che dallo spazio totale Stot = 20^N (nel “presupposto d’indipendenza”) Htagliato è passato subito al sottospazio Sa = 20^G, dove G < N proprio per tenere conto dei vincoli fisici, chimici, ecc.
Sarebbe bello avere da esperti come lei un ordine di grandezza di G…, come sollecitato dallo stesso articolista: se se ne ignora la richiesta, presentata con umiltà, la critica e l'ironia appaiono un po' preconcette, o no?
Legga bene: il problema originale posto da Htagliato non è tanto di probabilità, come sono soliti fare presso l'ID, ma piuttosto di un paradosso posto dall'intervallo delle estremità. Qualcosa da dire in proposito?
“Il problema di probabilità”, Nadia, è vero che non c’entra principalmente con l’articolo di Htagliato, però non è posto solo dall’ID – che come pars destruens assume spesso cmq posizioni scientificamente ineccepibili -, ma resta tuttora un macigno sulla strada del darwinismo anche fuori dall’ID. Vedi per es. Schützenberger o Kauffman.
Nadia, secondo me né Greylines né Nando hanno capito di cosa Htagliato stesse parlando: almeno è quel che se ne deduce leggendo i loro commenti.
Ma è sempre il rischio che si corre quando si è alla frontiera tra due discipline.
Ho letto il suo intervento e la proposta di simulazione e le chiedo: quali parametri dovrebbe avere tale simulazione?
Perchè, come in effetti già dissi tempo fa a Damasco, senza avere risposta, egli quando parla di sistema non omogeneo e stati di equilibrio (i.e. “attrattori”) mi ricorda da vicino una cosa che i fisici conoscono di solito, ovvero la dinamica molecolare.
Se modelliamo lo spazio “SA” come disomogeneo e non lineare, esso sarà caratterizzato da valli di stabilità separate da picchi di minore probabilità stanti i parametri.
La stabilità (ovvero la non necessità di cambiare) dipenderà dalla fitness verso l’ambiente, mentre i picchi di improbabilità rappresenteranno individui rari nella popolazione, meno adatti allo status quo ma potenzialmente più adatti ad un cambiamento, in caso di nuovi parametri.
Dopo che il sistema è condotto su un picco, da variazioni repentine dei parametri, verso quale valle scende, è definito da un evento caotico, ovvero una piccola variazione dei fattori può far rotolare l’ecosistema verso una valle o l’altra, dove il penso di diversi vincoli è diverso.
Per questo non capisco come lei possa dire che il “caso” (o meglio, l’effetto caotico) non abbia importanza, sopratutto quando pure Damasco pare essere un sostenitore delle oscillazioni, mentre in un tale modello avrebbe un ruolo determinante, anche di difficile predizione, a causa appunto della caoticità del sistema.
Credo che l’esercizio di HTagliato volesse “spezzare le gambe” al gradualismo Darwiniano e alla selezione naturale, non al caso in generale.
Gentile Nadia, mi permetta, il teatrino lo propone lei che arriva qui a fare il “carabiniere”, a dare patenti di professore, studente, umile ecc.
Il presupposto di indipendenza deriva dal numero astronomico E, messo a denominatore nella stima della probabilità. Un “vizio” quello di ignorare la conoscenza esistente che già riproverai a Giorgio Masiero per quel suo 20^200.
Come ho già detto Damasco non mostra la precisione e la dimestichezza verso gli argomenti di cui tratta quando stima l’effetto di eventuali vincoli come “più piccolo” e quando stima tale spazio come omogeneo. Abbastanza per far notare che un tale modello non può avere significato in merito ad un quesito fondamentale, ma che deve essere rivisto alla luce di tutti i dati se vuole avere tale obiettivo.
La domanda “questo numero G di proteine possibile in base ai vincoli che numero è?” è una domanda a cui sarà possibile rispondere quando e se mai avremo capito come le proteine si sono evolute.
In tal senso ho semplicemente fatto notare a Damasco che se vuole modellare il problema sarebbe opportuno lo facesse partendo da tutto quello che si sa a riguardo, per poter capire qualcosa in più.
Il “numero astronomico” E può impressionare solo lei, Nando. Io l’ho imparato leggendo l’Orologiaio cieco del darwinista Dawkins, che lo calcolava dall’emoglobina e lo usava, senta senta, per dimostrare il funzionamento intelligente del setaccio della selezione naturale.
Htagliato invece (come Masiero a suo tempo, in altro articolo) lo usa come massimante per dimostrare l’esistenza di “vincoli” fisico-chimici, che ancora non conosciamo, lei mi conferma, così da arrivare al numero G.
Insomma
1) la sua dotta lezione a Greylines non c’entra nulla con quanto Htagliato afferma in questo articolo;
2) lei ha confuso un massimante matematico con un calcolo di probabilità;
3) non conosce un massimante minore di E.
Forse Htagliato conosce bene la fisica e la matematica e sa poco di biologia, ma almeno si fa aiutare per i suoi articoli peer review di biologia evolutiva da un biologo. Lei da chi si fa aiutare per la matematica?
Temo Nadia che lei non sia molto interessata al dialogo, quanto piuttosto a lanciare accuse. Lungi da me la volontà di contrariarla, constatato quanto sopra, la lascio di buon grado della sua opinione, soprattutto alla luce di come l’ha argomentata e di come io abbia già scritto quanto sufficiente a risponderle.
Il ragionamento di HTagliato mi sembra chiaro che sia stato volutamente impostato su delle astrazioni concettuali per ridurre al minimo indispensabile la parte computazionale proprio perchè il calcolo realistico delle effettive probabilità non sarebbe necessario visto che HTagliato lo rende inutile dalle implicazioni logiche del ragionamento stesso…
Bello,bellissimo rivedere la firma di un “grande”:Htagliato.Simpatico e modesto.
Bella riflessione che potrebbe essere lo spunto iniziale di una ricerca interessante.
Ovviamente la seconda ipotesi sembra essere quella intelletteualmente la più soddisfacente nel quadro delle ipotesi da te poste: densità STOT non uniformi causati dai vincoli che a priori favoriscono la fitness. Varrebbe la pena confermare quest’intuizione con una serie di simulazioni.
Detto ciò, epistemologicamente, non sono sicuro che ci sia una così grande differenza tra l’ipotesi 1 e l’ipotesi 2 nel senso che nei due casi vi è de facto distruzione dello spazio di fase raggiungibile e quindi la susseguente perdita di informazione essendo significativa.
Ad ogni modo e qualunque sia il modello, il ruolo del “caso” , “inteso come meccanismo indipendente dalla fitness” come dici benissimo è ridotto vicinissimo a zero, solo i vincoli statisticamente influenti conducendo alla la riduzione dello spazio delle fasi e, quindi, alla nuova struttura vi sono significativi.
CHIARIMENTI SULL’ARTICOLO
Cari lettori di Critica Scientifica,
mi dispiace molto non avere più il tempo libero di una volta, per cui non scrivo e non intervengo con la frequenza di un tempo. Lo faccio in questa occasione perché l’articolo in esame espone calcoli nuovi e non facilmente fruibili, come si evince dalla grande confusione emersa in alcuni commenti tra ieri e oggi.
Innanzitutto, operiamo una sintesi: questi calcoli NON sono calcoli di probabilità ma riguardano condizioni da rispettare PRIMA di un eventuale calcolo di probabilità, sono propedeutici all’avere una probabilità che non sia nulla. L’interdipendenza delle reazioni quindi non interviene ancora (interverrebbe se volessimo passare da una probabilità semplice ad una composta), mentre è assurdo obbiettare che non si è tenuto conto dei vincoli, significherebbe non aver letto l’articolo.
Il senso dell’articolo è il seguente: da un punto di vista matematico, possono convivere nella stessa teoria l’assenza di gradualismo e un’impostazione di tipo neodarwiniano, basata su una sorgente di cambiamenti indipendenti dalla fitness regolarizzata dalla selezione naturale? La risposta breve è NO. La risposta lunga è che o assumo la prima condizione o l’altra ma non entrambe contemporaneamente, altrimenti dovrei snaturare la definizione stessa di meccanismo neodarwiniano. La risposta completa coincide con l’articolo.
Ora entriamo in alcuni dettagli importanti. Per quanto riguarda i VINCOLI, nel modello li ho resi estremamente severi, dicendo che il loro effetto è di BLOCCARE del tutto un aminoacido, per cui da che ne sono N è come se ne fossero solo G. Non credo che potevo renderli più severi di così, ma accetto proposte in tal direzione.
ATTENZIONE alla funzione esponenziale, che non è per niente intuitiva: lo spazio delle fasi accessibile NON è “un po’ più piccolo” di quello totale, perché ogni volta che introduco un vincolo definito come sopra, tale spazio si riduce di un fattore 20 (è un ventesimo di quello precedente, immediatamente più largo); per es. se con un solo vincolo lo spazio accessibile è il 5% di quello iniziale, con due vincoli diventa il 0,25%, con tre vincoli il 0,0125%, con quattro il 0,000625% e così via. Dire che la riduzione dello spazio sia stata drastica è dire poco.
Smontiamo subito invece l’obiezione delle reazioni in serie anziché in parallelo: se un intervallo di tempo (per es. un’era geologica) lo divido per la durata della più piccola delle trasformazioni fisico-chimiche, ottengo il numero massimo delle reazioni che può aver compiuto nel tempo a disposizione un solo atomo, quindi, moltiplicando tale numero per il numero di atomi, ottengo tutte le reazioni compiute da tutti gli atomi in quel periodo (quindi, in “parallelo”). Se inserissi qui un’interdipendenza (del tipo se l’atomo A reagisce con B, B sta reagendo con A per cui ho una reazione e non due) avrei meno reazioni possibili, andando ancora più severamente contro la realizzabilità del neodarwinismo e non a favore!
Passando alla seconda parte dell’articolo, sono invece lieto che siamo tutti d’accordo che la prima ipotesi (densità delle proteine potenzialmente realizzabili uniforme) sia stata rigettata da tutti. Ottimo! Sono cose che gli esperti in proteine già sanno.
Il vero problema sorge che non posso abbracciare la seconda ipotesi (densità non uniforme, ma anzi concentrata sui punti a massima fitness delle proteine già esistenti che fanno bene il loro lavoro) senza accettare le palesi conseguenze. O meglio, diventa un problema di coerenza, del tipo “botte piena VS moglie ubriaca”: alcuni vorrebbero contemporaneamente vincoli severi ma non così tanto da non lasciare più spazio al meccanismo “variazione indipendente dalla fitness + selezione” di agire. Se si vogliono entrambe le cose, allora occorre fare dei calcoli, non discorsi sulla plausibilità.
Qui purtroppo siamo costretti a ribadire l’ovvio: la selezione naturale agisce solo DOPO una qualsiasi mutazione, non prima, perché non stiamo in un’azienda dove il prodotto X non viene più prodotto se non c’è nessuno che lo compra. La selezione può indirizzare all’interno della popolazione già esistente, non nello spazio costruito a priori di cui si parla nell’articolo.
Se quindi i vincoli dipendono dalla fitness, cade meno la prima condizione DEFINITORIA di neodarwinismo. Infine, il tipo di stabilità che i vincoli vanno a creare è una stabilità A PRIORI, è come un attrattore che non dipende dal tempo che fa, la specie non vi ci sta dentro per ragioni di contingenza. La selezione naturale, quella sì, è per definizione contingente (non esistendo un criterio universale a priori per stabilire quale sia il fenotipo più adatto), ma la selezione è inutile se il tipo di variazioni su cui interviene non sono indipendenti dalla fitness, perché allora il merito dell’evoluzione va a queste e non alla selezione.
Riassumendo: gradualismo NO (e siamo d’accordo), vincoli a priori legati alla stabilità e quindi in parte alla fitness Sì (e siamo d’accordo), spazio delle fasi ridotto da un universo di possibilità ad un granello di polvere Sì (e siamo d’accordo), come lo vogliamo chiamare questo? Neodarwinismo? Se sì, non sono d’accordo, ma fate come volete, “What’s in a name….?”, diceva Shakespeare.
Elementare, Watson…, ma guai a toccargli Darwin!
Gentile Damasco,
discutere di A e B dicendo che non si tratta di un calcolo di probabilità e poi dire che A è il numero di casi d’interesse e B il numero di casi possibili mi pare poco coerente. Lei ha fatto una discussione trattando elementi che possono comporre una valutazione di probabilità e di tali elementi si è discusso. Lei ha parlato di “200” sarebbe stato più opportuno un numero inferiore, meglio ancora non dare numeri quando non se ne conosce l’entità.
In merito a tali elementi si è indicato come in effetti non sia opportuno definirne uno come composto da eventi indipendenti e si tratta di una questione molto nota in biologia, ovvero di come in una evoluzione, per definizione, il passo precedente condiziona quello successivo.
Il suo tentativo “matematico” di tranciare una discussione sulla base di premesse tanto confuse mi pare poi francamente avventato. Lei non pare avere dimestichezza con la biologia, come le ho fatto notare gli amminoacidi in una proteina non sono casuali. Ad esempio, se ha un triptofano sarà molto difficile che ne abbiamo un altro successivamente. Le mutazioni che fanno passare da un AA ad un altro non sono casuali, passare da un codone che codifica per P ad uno che codifica per S non è come passare ad uno che codifica per T (le lascio la ricerca della ragione come stimolo ad approfondire) e ci sono molti altri vincoli spaziali, ricordi che le proteine sono espresse a partire dal DNA e su di esso altri vincoli devono essere valutati (accessibilità, cromatina, ecc). Modellare semplicemente la sequenza aminaocidica come una serie di eventi e mettere tutto esponente 20 è la classica mucca approssimata con una sfera; una storiella sui fisici più ingenui che piace molto ai biologi.
Lo spazio delle proteine adatte è, come già detto, uno spazio non lineare che può immaginare come un iperspazio fatto di picchi e valli di possibili configurazioni stabili e funzionali della proteina. Tale spazio è influenzato da tutte le molecole con cui la proteina e il suo codice genetico entrano in contatto, ponendo la necessità di un modello molto complesso di quello da lei delineato.
Un evento selettivo avrà la funzione di far muovere la proteina da un punto di equilibrio ad un altro e questo sarà condizionante rispetto alla possibilità di evolvere ulteriormente. Lo spazio può variare gradualmente come variano tutti i fattori influenti la fitness ed eventi drastici, sia molecolari (la delezione di un gene) che ambientali (un repentino cambiamento ambientale) avranno la possibilità di configurare nuovi eventi. Non pare un ipercristallo omogeneo e regolare che risuona, ma piuttosto un iperpaesaggio con diverse densità e caratteristiche iperorografiche a influenzare i percorsi.
Noti ad esempio che viviamo un uno spazio in cui gli AA sono enantiomeri di tipo L, come la mano sinistra. Gli speculari D avrebbero potuto fare esattamente lo stesso compito in un mondo “a specchio” del nostro. Cosa a spinto la biologia dal lato L piuttosto che da quello D?
Lei spinge per un vincolo pregresso e sta a lei dire quale, altri non hanno lo stesso onere e rispondono con un evento caotico, selettivo, su cui non è quindi utile soffermarsi, data la sua instabilità. Infatti poteva andare in un altro modo, ma non è stato così, a causa di uno dei numerosi fattori epsilon, che hanno operato una prima selezione in un mondo un cui gli AA sono sia L che D.
Qual è la relazione tra vincoli e fitness? Se una proteina si trova in uno punto dell’iperspazio non è ragionevole pensare che ci sia arrivata “saltando” più volte, ma seguendo il percorso per certi “iperpassi” tra ipercanyon tra iperpicchi, con pochi “salti” legati a pochi eventi drastici. Perchè i codoni sono fatti ad un certo modo, perchè la cromatina è accessibile in un certo modo, e possibilmente molte altre ragioni di biologia di base che non conosciamo e potrebbero essere essenziali a capire, a modellare il fenomeno (si immagini lo stesso esercizio prima di Watson & Crick..).
Veda a tal proposito ad esempio il lavoro di S. Gavrilets, Sergey. “Evolution and speciation in a hyperspace: the roles of neutrality, selection, mutation and random drift.” In Towards a comprehensive dynamics of evolution-exploring the interplay of selection, neutrality, accident, and function. Oxford Univ. Press, New York del 2003
La ragione di un certo ipercammino non è del tutto casuale, ma si basa su eventi pregressi, che hanno indotto dei vincoli (L piuttosto che D, relazioni tra i codoni, la fusione con un dominio in uno pseudogene, ecc.). Tale percorso non è necessariamente “un aminoacido alla volta”, come accennato, nel neodrwinismo c’è anche la ricombinazione, di cui lei non tiene conto.
Il DNA non si ricombina una base alla volta, ma a blocchi e specie diverse hanno ordine nettamente diverso anche in relazione a geni ortologi. Il modello che lei propone non tiene conto della complessità e della conoscenza del fenomeno biologico di cui cerca di modellare l’evoluzione e quindi non è per esso significativo, come la mucca sferica non è significativa per la mucca.
Tutto l’iperspazio di cui lei tratta (e ha ipotizzato persino omogeneo!) è poi dinamico non lineare, non statico e dipende da eventi esterni (temperatura, umidità, altre specie, altre proteine, altri geni, pseudogeni, ecc. ecc.).
Tale iperspazio appare inoltre caotico. Commetta un errore anche piccolo e il modello non sarà utile, divergendo rapidamente e significativamente dal fenomeno caotico reale.
Vuole calcoli dice. Li ha cercati?
In letteratura ne trova di molto approfonditi e in grande quantità. Riguardo alla caoticità di taluni domini proteici, può ad esempio consultare il lavoro di VN Uversky “Dancing protein clouds: the strange biology and chaotic physics of intrinsically disordered proteins.” su JBC del 2016. Segua i riferimenti nell’articolo, troverà lavori che trattano di modellazione.
Riassumendo: gradualismo POSSIBILE; non vuol dire un aminoacido alla volta, ma un cambiamento alla volta, che non è detto sia su un aminoacido solo (ricordi che alla base sta il DNA). Vincoli dinamici, non indipendenti dal pregresso ipercammino, in un iperspazio dinamico e caotico. Eventi caotici, con possibilità di far cambiare drasticamente l’ipercammino.
Come vuole chiamare un percorso che dipende dal suo passato, in cui un singolo evento può cambiare in modo netto il destino delle generazioni successive, decretando la fine di altre che si sono avviate per un evento epsilon verso un ramo dell’ipercanyon diverso, che può anche cambiare di pendenza per effetto del cammino di altri su altri rami dell’ipercanyon stesso?
Come le ho già scritto, penso che il suo giovane intelletto sarebbe messo a migliore frutto leggendo un po’ di più su quanto si sa (un dottorato in biologia evolutiva?) e su quanto lei vuol normare definitivamente. Per dedurre sulla base di nozioni più solide e concrete, rispetto alla visione “sferica” che ha del fenomeno su cui cerca così tenacemente di mettere punti definitivi.
Grazie Nando per questa sua bellissima pagina, in cui, per chi fosse veramente interessato ad analizzarla profondamente, si possono cogliere molti degli elementi epistemologici fondamentali che distinguono la biologia, in particolare la biologia evoluzionistica, dalla fisica, elementi che nel mio piccolo ho sempre tentato, con tutti i miei limiti, di divulgare nei miei articoletti: storicità, complessità, emergenza, principio di indifferenza. Ed è proprio a quanto da lei magistralmente spiegato che mi riferivo nel mio commento quando manifestavo la mia perplessità nel fatto di non riuscire a cogliere, nel pur interessante articolo di Htagliato, la necessaria linea di continuità con il quadro storico-biologico generale.
Mah, io dalla tiritera di Nando deduco che non ha capito nulla del ragionamento di Htagliato. Per cominciare, Lei, Vomiero, mi saprebbe spiegare perché Htagliato avrebbe dovuto scegliere (secondo Nando) un numero di residui minore di 200?
Come scrissi ieri (ricorda che le dissi che avevo già risposto ai suoi quesiti?)
“Per cominciare un appunto sul concetto che lei da di proteina “semplicissima”. La invito a consultare un lavoro sulla lunghezza media di una proteina, in particolare di quelle più evoluzionisticamente “recenti”, come ad esempio quello pubblicato su Genome Biology qualche anno fa (2010) da Capra JA et al. intitolato “Novel genes exhibit distinct patterns of function acquisition and network integration” 200AA è una lunghezza media per una proteina “recente”, con il 25esimo percentile più intorno a 100AA (proteine “corte”) e quelle “molto corte” (semplicissime come dice lei), nettamente sotto ai 100AA”
Continuo a non capire, Nando, perché lei rimprovera ad Htagliato la scelta di 200 aa. Non capisco per 2 motivi: il principale che tra 100 e 200 nulla cambia del “paradosso” logico di Htagliato, il secondario che – comunque – gli studi danno per la lunghezza media delle proteine degli eucarioti 400 aa e dei procarioti 300 aa.
Damasco nel suo modello cerca di trattare di proteine “semplicissime” e non considerando fenomeni di ricombinazione e fusione. Tali proteine non sono “medie” (che son lungi dall’essere semplici), ma piuttosto recenti e brevi, come indicato nel lavoro che ho citato (lo ha scorso?).
Siamo del tutto d’accordo che ai fini del modello non cambia nulla, ma se Damasco decide un numero, non lo deve fare “a naso” oppure può benissimo (come consigliato) mettere un valore “x” e dire “piccolo relativamente alla taglia della proteina” e fine della discussione.
Viceversa, il problema di quel “200” è che poi si trovano testi come quelli di Masiero dove il dato è preso alla lettera ed elevato alla 20, quindi essere precisi è importante.
Non sono d’accordo. Il numero 200 non l’ha scelto né Htagliato né Masiero, ma è il numero tipico usato in letteratura scientifica quando ci si approccia alla stocastica della sintesi proteica. Le ho già citato il pioniere Dawkins con riferimento all’emoglobina nel suo Orologiaio cieco, Le aggiungo Stuart Kauffman di Esplorazioni evolutive (esattamente alla pag. 189 nella mia Edizioni Einaudi del 2005), il quale parte proprio dalla lunghezza 200.
“Essere precisi è importante”, conclude lei. Htagliato è stato preciso.
Quella non è una proteina “semplicissima”.
Io non credo che il problema sia il numero di AA per la proteina standard considerata, Nadia. Credo piuttosto sia una questione di approccio. Nell’articolo di Htagliato, che ripeto, trovo comunque interessante, vedo che si parla di proteine senza quasi mai fare riferimento né agli step biologici che servono per avviare il processo, cromatina, DNA, RNA messaggero, RNA di trasporto, ribosoma, codoni ecc., né a quelli che lo completano, folding proteico. Ora, forse in fisica l’approccio riduzionistico potrà anche essere di grande utilità, isolo il mio sistema, lo rendo ideale, lo studio, lo modellizzo, e vedo cosa succede. In biologia purtroppo non è quasi mai così, ci sono processi fortemente connessi tra di loro, ci sono sistemi complessi in continua interazione tra di loro e con l’ambiente, che si ristrutturano e cambiano la loro organizzazione interna rendendosi così particolarmente resistenti alla modellazione fisico-matematica. In definitiva quindi, concordo con Nando sul fatto che il modello proposto da Htagliato è probabilmente ipersemplificato e di conseguenza temo che possa anche essere di scarsa utilità pratica per il biologo.
Lei non crede, Vomiero, che la lunghezza scelta da Htagliato sia importante, Nando lo crede invece importante. Su un punto del modello di Htagliato lei e Nando avete idee opposte.
Lei si sbaglia Nadia, l’importante è la considerazione per gli aspetti biologici e quando Damasco dice che una proteina da 200AA è “semplicissima” fa capire quanto la cosa lo interessi. Gli si è fatto notare che se vuole che qualcuno prenda il suo modello sul serio, deve fare uno sforzo di realismo maggiore, oltre che essere più preciso. Come già detto e ripetuto, il 200 nel modello non conta, poteva benissimo essere un “x” generico (i problemi sono altri, come evidenziato), ma conta nella quota di precisione, tanto che poi quel numero rischia di diventare seriamente al centro di un modello e poi emergono altri problemi. Una proteina semplicissima non è in media di 200AA, questo è un fatto.
“Il 200 nel modello non conta”, lei ammette, però poi ne fa un caso soffermandosi al significato tecnico dell’aggettivo “semplicissima”, quando in questo articolo divulgativo Htagliato ha usato l’aggettivo nel senso comune, per sottintendere che ci sono proteine anche molto più lunghe… da 1.000 o da 10.000 aa, sappiamo. Non ho altro da aggiungere sulla questione.
Io dico che si sbaglia lei, lei dice che mi sbaglio io. Chi ci legge deciderà per conto suo.
Come già le dissi Nadia, non ho alcun interesse nel contrariarla. Se lei interpreta le cose così, sta bene.
A me pare, che c’è proprio un’incomprensione di base dell’articolo di Htagliato: leggendolo mi pare che lui stia considerando un insieme microcanonico dal punto di vista statistico (che è un discorso aldilà del probabilistico).
Mentre, leggendo Nando porsi la domanda “Qual è la relazione tra vincoli e fitness? “ , io vi leggo una preoccupazione neanche canonica ma proprio macrocanonica.
In altre parole per rispondere a quest’ultima domanda bisognerebbe già ipotizzare una legge di moto nello spazio di fase che descrivesse la dinamica del sistema considerato (e che, sola, sarebbe atta a potenzialmente risolvere la problematica legata ai “picchi”, “passi” e altri “salti”…)
Non penso che era la problematica di Htagliato e quindi questo commento sà un poco di cavoli alla merenda.
Comunque mi posso sempre sbagliare ; ne sapremo di più quando Htagliato avrà tempo di rispondere.
Il mio argomento è che il modello di Damasco sia ipersemplicistico. Ho un backgroud in fisica modesto, ma capisco la definizione di microcanonico, canonico e gran canonico.
A conferma della mia osservazione e ad esemplificare un approccio analogo a quello di Damasco ma più approfondito, questo lavoro:
P Davies et al “Self-organization and entropy reduction in a living cell.” in BioSystems del 2013
conferma che un modello canonico sarebbe da preferire, in virtù appunto della diversa frequenza degli AA e che anzi, essendo il sistema aperto (e il lavoro è focalizzato sulla cellula, mentre noi parliamo di ecosistema) sarebbe da preferire un modello gran canonico.
Se si vuole percorrere la strada di una valutazione entropica per modellare gli eventi biologici e fare considerazione di selezione naturale, un lavoro interessante potrebbe essere questo:
P Davies et al “Implications of quantum metabolism and natural selection for the origin of cancer cells and tumor progression.” AIP advances 2012
In ogni caso, come ho già fatto notare a Damasco, di lavoro analogo a quello che abbozza qui, ce n’è molto e di alta qualità e sarebbe opportuno che lo consultasse prima, per così dire, di accendere il fornello per scaldare l’acqua di un ennesimo modello termodimanico.
Siamo poi d’accordo che un tale iperspazio, ammesso sia sufficientemente preciso, permetterebbe assieme ad una opportuna legge di moto, di trovare una (e non “la”) possibile via che terminerebbe al punto in cui una proteina si trova oggi, fermo restando pure che non si sa da dove sia partita.
Giova prima di cimentarsi in tale impresa, notare che per la “mera dinamica” molecolare, si dispone di efficaci formule per simulare il fenomeno a livello quantistico e che ciò teoricamente condurrebbe ad una realista simulazione del moto della proteina.
Salvo il fatto che pure il più potente dei super calcolatori, con un peptide di qualche aminoacido, nel vuoto, impiega un tempo notevole per simulare qualche millisecondo di dinamica e che la complessità è esponenziale al numero di atomi del sistema. Ci si trova dunque con sistemi “a molle” che considerati i modesti risultati nel contesto del folding proteico, hanno ancora un bel po’ di sviluppo davanti prima di essere utili.
I limiti della modellazione paiono quindi piuttosto evidenti e la necessità di capire meglio la biologia per trovare una “scorciatoia” ovvero un’efficace livello di astrazione non è una via negoziabile a mio avviso, considerata la massa di cose che non sappiamo e sui cui ci poniamo quesiti.
Oppure assumiamo che la mucca sia sferica, ma poi ricordiamoci che la realtà là fuori è un’altra cosa.
Certo, Nando, il fenomemeno evoluzionistico è da capirsi come un sistema termodinamico aperto e bisogna applicarvi il secondo principio di termodinamica.
Ti consiglio la lettura di quest’articolo assolutamente appassionante, circa il quale, penso che te ed io potremmo incontrarci (intellettualmente parlando):
Natural process – Natural selection, Vivek Sharma, Arto Annila Biophysical Chemistry 127 (2007) 123–128
http://www.helsinki.fi/~aannila/arto/natprocess.pdf
“Abstract
Life is supported by a myriad of chemical reactions. To describe the overall process we have formulated entropy for an open system undergoing chemical reactions. The entropy formula allows us to recognize various ways for the system to move towards more probable states.
These correspond to the basic processes of life i.e. proliferation, differentiation, expansion, energy intake, adaptation and maturation. We propose that the rate of entropy production by various mechanisms is the fitness criterion of natural selection. The quest for more probable states results in organization of matter in functional hierarchies.”
Quanto alle mucche sferiche non le guarderei con disprezzo finché ti limiti a considerazioni in relazione al volume e fintanto che la forma stessa del bovino non sia importante per le considerazioni che ti interessano….
Ho spiluccato l’articolo ed è in effetti molto interessante. Grazie per la segnalazione!